
Sonke sisengabantwana safundiswa ukuba kukho izivamvo ezi-5: ukubona, ukungcamla, isandi, ukujoja, nokuchukumisa. Izivamvo ezine zokuqala zisebenzisa amalungu acacileyo, ahlukileyo, njengamehlo, amaqhuqhuva okungcamla, iindlebe, impumlo, kodwa imvakalelo yomzimba ichukumisa njani kanye? Ukuchukumisa kubonakala kuwo wonke umzimba, ngaphakathi nangaphandle. Akukho lungu elinye elahlukileyo elinoxanduva lokuva ukubamba. Endaweni yoko, kukho ii-receptors ezincinci, okanye i-nerve endings, ejikeleze umzimba wonke eziva uchukumisa apho yenzeka khona kwaye ithumela imiqondiso ebuchotsheni ngolwazi malunga nohlobo lokuchukumisa okwenzekileyo. Njengoko i-taste bud ngolwimi ibona i-flavour, i-mechanoreceptors i-gland ngaphakathi kwesikhumba kunye nezinye izitho ezibona ukuvakalelwa. Baziwa ngokuba i-mechanoreceptors kuba ziyilelwe ukufumanisa iimvakalelo zoomatshini okanye umahluko kuxinzelelo.
Contents
Indima yeeMechanoreceptors
Umntu uyaqonda ukuba baye bafumana imvakalelo emva kokuba ilungu elijongene nokufumanisa loo ngqiqo ethile ithumela umyalezo kwingqondo, eyona nxalenye ephambili eyenza kwaye ilungiselele yonke ingcaciso. Imiyalezo ithunyelwa ukusuka kuzo zonke iindawo zomzimba isiya ebuchotsheni ngeengcingo ezibizwa ngokuba ziineurons. Kukho amawaka ee-neuron ezincinci eziphuma kuzo zonke iindawo zomzimba womntu, kwaye kwiziphelo ezininzi zezi neurons zii-mechanoreceptors. Ukubonisa okwenzekayo xa uchukumisa into, siya kusebenzisa umzekelo.
Yiba nombono wengcongconi uhlala engalweni yakho. Ubunzima bale nambuzane, ukukhanya kakhulu, kuvuselela i-mechanoreceptors kuloo ndawo ethile yengalo. Ezo mechanoreceptors zithumela umyalezo ecaleni kwe-neuron eziqhagamshelwe kuyo. I-neuron idibanisa yonke indlela eya kwingqondo, efumana umyalezo wokuba kukho into echukumisa umzimba wakho kwindawo echanekileyo ye-mechanoreceptor ethile ethumele umyalezo. Ingqondo iya kwenza ngeli cebiso. Mhlawumbi iya kuxelela amehlo ukuba ajonge kwingingqi yengalo efumene utyikityo. Kwaye xa amehlo exelela ingqondo ukuba kukho ingcongconi engalweni, ingqondo inokuxelela isandla ukuba siyiphephethe ngokukhawuleza. Yileyo ndlela i-mechanoreceptors isebenza ngayo. Injongo yale nqaku ingezantsi kukubonisa kunye nokuxoxa ngokweenkcukacha umbutho osebenzayo kunye ne-molecular determinants of mechanoreceptors.
I-Touch Sense: Umbutho oSebenzayo kunye ne-Molecular Determinants ye-Mechanosensitive Receptors
Abstract
I-mechanoreceptors ye-cutaneous ifakwe kwiindawo ezahlukeneyo zesikhumba apho zibona uluhlu olubanzi lwe-stimuli yomatshini, kubandakanywa ibrashi yokukhanya, i-stretch, i-vibration kunye noxinzelelo oluyingozi. Olu luhlu lwe-stimuli luhambelana noluhlu oluhlukeneyo lwee-mechanoreceptors ezikhethekileyo ezisabela kwi-deformation ye-cutaneous ngendlela ethile kwaye idlulisele ezi zikhuthazo kwizakhiwo eziphezulu zengqondo. Izifundo kuzo zonke ii-mechanoreceptors kunye neziphelo zemithambo-luvo ezinofuzo ziqala ukufumanisa iindlela zokuchukumisa. Umsebenzi kule ntsimi unike abaphandi ukuqonda ngokucokisekileyo ngakumbi kwintlangano yesiphaluka ephantsi kombono wokuchukumisa. Imijelo ye-ion entsha iye yavela njengabaviwa beemolekyuli zokutshintshwa kunye neepropathi zemisinga ye-mechanical gated yaphucula ukuqonda kwethu iindlela zokuziqhelanisa ne-tactile stimuli. Olu phononongo lubonisa inkqubela phambili eyenziwe ekubonakaliseni iimpawu ezisebenzayo ze-mechanoreceptors kwizinwele ezinoboya kunye ne-glabrous kunye nemijelo ye-ion ebona igalelo lomatshini kunye nokulungiswa kwe-mechanoreceptor.
Internet: i-mechanoreceptor, i-chanosensitive channel, intlungu, isikhumba, inkqubo ye-somatosensory, ukubamba
intshayelelo
Ukuchukumisa kukufumanisa isivuseleli soomatshini esichaphazela ulusu, kubandakanywa nefuthe elingenabungozi kunye neliyingozi. Yingqiqo ebalulekileyo kubomi kunye nophuhliso lwezilwanyana ezanyisayo kunye nabantu. Uqhagamshelwano lwezinto eziqinileyo kunye nolwelo olulusu lunika ulwazi oluyimfuneko kwinkqubo ye-nervous central evumela ukuhlola kunye nokuqatshelwa kokusingqongileyo kwaye iqalise ukuhamba okanye ukunyakaza kwesandla okucwangcisiweyo. Ukuchukumisa kubaluleke kakhulu ekuqeqesheni umsebenzi, ukunxibelelana nabantu kunye nokwabelana ngesondo. Uluvo lokuthintela lulona luvo lubuthathaka, nangona lunokuphazamiseka (hyperesthesia, hypoesthesia) kwiimeko ezininzi ze-pathological.1-3
Iimpendulo zochuku zibandakanya ikhowudi echanekileyo yolwazi lomatshini. I-mechanoreceptors ye-Cutaneous ifakwe kwiindawo ezahlukeneyo zesikhumba apho zibona uluhlu olubanzi lwe-stimuli yomatshini, kubandakanywa ibrashi yokukhanya, ukunweba, ukungcangcazela, ukuphambuka kweenwele kunye noxinzelelo oluyingozi. Olu luhlu lwe-stimuli luhambelana noluhlu oluhlukeneyo lwee-mechanoreceptors ezikhethekileyo ezisabela kwi-deformation ye-cutaneous ngendlela ethile kwaye idlulisele ezi zikhuthazo kwizakhiwo eziphezulu zengqondo. I-Somatosensory neurones yesikhumba iwela kumaqela amabini: i-mechanoreceptors ephantsi (i-LTMRs) esabela kwingcinezelo ye-benign kunye ne-high-threshold mechanoreceptors (HTMRs) ephendula ukuvuselela umatshini onobungozi. Iiseli ze-LTMR kunye ne-HTMR zihlala ngaphakathi kwe-dorsal root ganglia (DRG) kunye ne-cranial sensory ganglia (i-trigeminal ganglia). Iifayibha zeNerve ezinxulumene ne-LTMRs kunye ne-HTMRs zihlelwa njenge-A?-, A?- okanye i-C-fibers ngokusekelwe kwizenzo zabo ezinokuthi ziqhube ngokukhawuleza. Iintsinga ze-C azinamyelinated kwaye zinezona santya zicothayo zokuqhuba (~2 m/s), kanti A? kunye no-A? iintsinga zikhaphukhaphu kwaye zininzi i-myelinated, zibonisa eziphakathi (~12 m/s) kunye nesantya sokuqhuba esikhawulezayo (~20 m/s), ngokulandelelanayo. Ii-LTMR zikwahlulahlulwe ngokukhawuleza, okanye ngokukhawuleza ukulungelelanisa iimpendulo (i-SA- kunye ne-RA-LTMRs) ngokwemilinganiselo yazo yokulungelelanisa kwi-stimulus eqhubekayo yomatshini. Ziyakwahlulwa ngakumbi ngamalungu okuphela kwesikhumba angenasiphelo kunye nezivuseleli ezizithandayo.
Ikhono le-mechanoreceptors lokubona iimpawu zomatshini zixhomekeke ebukhoneni beendlela ze-ion ze-mechanotransducer eziguqula ngokukhawuleza amandla omatshini kwiimpawu zombane kunye nokulahla intsimi eyamkelayo. Oku kuchithwa kwendawo, okubizwa ngokuba yi-receptor potential, kunokuvelisa izinto ezinokubakho ezisasaza kwinkqubo ye-nervous central. Nangona kunjalo, iipropathi zeamolekyu ezidibanisa i-mechanotransduction kunye nokulungelelaniswa kumandla omatshini zihlala zingacacanga.
Kolu hlaziyo, sinika umboniso weempawu ze-mammalian mechanoreceptor kwi-touch engabonakaliyo kunye neyingozi kwisikhumba esinoboya kunye ne-glabrous. Sikwaqwalasela ulwazi lwakutsha nje malunga neepropathi zemisinga ye-mechanically-gated currents kwinzame yokuchaza indlela yokulungiswa kwe-mechanoreceptor. Ekugqibeleni, sihlaziya inkqubela phambili yakutshanje eyenziwe ekuchongeni iziteshi ze-ion kunye neeprotheni ezihambelanayo ezijongene nokuveliswa kwee-currents ze-mechano-gated.
Innocuous Touch
I-Hair Follicle-Associated LTMRs
Iinwele zeenwele zimele i-hair shaft-produceing mini-organs ezibona ukukhanya. Iifayibha ezinxulumene neenwele zeenwele ziphendula ukunyakaza kweenwele kunye nolwalathiso lwazo ngokudubula oololiwe bezinto ezinokubakho ekuqaleni kunye nokususwa kwe-stimulus. Bazivumelanisa ngokukhawuleza ii-receptors.
Ikati kunye nomvundla. Kwingubo yekati kunye nomvundla, iifolli zeenwele zinokuhlulwa zibe ziintlobo ezintathu zeenwele zeenwele, iNwele ePhantsi, inwele yoMgada kunye neTylotrichs. Iinwele ezisezantsi (iinwele ezingaphantsi, uboya, i-vellus)4 zezona zininzi, ezona zinwele zimfutshane kunye nezona zintle zedyasi. Zingamaza, azinambala kwaye zavela ngokwamaqela eenwele ezimbini ukuya kwezine ukusuka kwindawo eqhelekileyo eluswini. Iinwele zoonogada (ii-monotrichs, overhears, tophair)4 zigobile kancinane, mhlawumbi zinebala elibomvu okanye azinabala, kwaye zivele zodwa emilonyeni yeefollicle zazo. I-tylotrichs yizona zincinci kakhulu, iinwele ezide kunye nezona zide kakhulu. Iifiber zenzwa zokubonelela kwi-follicle yeenwele zifumaneka ngaphantsi kwe-sebaceous gland kwaye zibangelwa ngu-A? okanye A?-LTMR imicu.5,6
Kwindawo esondeleyo kwi-shaft yeenwele ezisezantsi, ngaphantsi nje kwinqanaba le-sebaceous gland yi-ring of lanceolate pilo-Ruffini endings. Ezi ziphelo zemithambo-luvo zibekwe kwikhosi ejikelezayo ejikeleze i-shaft yeenwele ngaphakathi kwethishu edibeneyo eyenza i-follicle yeenwele. Ngaphakathi kwi-follicle yeenwele, kukho iziphelo ze-nerve zamahhala, ezinye zazo zenza i-mechanoreceptors. Ngokuqhelekileyo, ii-corpuscles ezithintekayo (bona ulusu olukhanyayo) zijikeleze intamo ye-tylotrich follicle.
Iipropathi ze-myelinated nerve endings kwikati kunye ne-rabbit hairy skin ziye zaphononongwa ngokugqithiseleyo kwixesha le-1930-1970 (uphononongo ku-Hamann, 1995) kunye nomvundla, baye bahlula iimpendulo kwiintlobo ezintathu ze-receptor ezihambelana nokunyakaza kweenwele eziphantsi (uhlobo lwe-D receptors), Iinwele ze-Guard (uhlobo lwe-G receptors) kunye neenwele zeTylotrich (uhlobo lwe-T receptor) .8 Zonke iimpendulo ze-nerve fiber zihlanganiswe ndawonye. kwi-Rapidly Adapting receptor yohlobo lwe-I (RA I) ngokuchaswa kwe-Pacinian receptor ebizwa ngokuba yi-RA II. I-RA I-mechanoreceptors ibona i-velocity ye-stimulus mechanical kwaye inomda obukhali. Ababoni ukuhluka kwe-thermal. Burgess et al. iphinde yachaza i-receptor yentsimi eguqukayo ngokukhawuleza ephendula ngokufanelekileyo ekubetheni kwesikhumba okanye ukuhamba kweenwele ezininzi, eziye zabangelwa ukuvuselela iziphelo ze-pilo-Ruffini. Akukho nanye impendulo ye-follicle yeenwele ezibangelwa ngumsebenzi we-C fiber.772
Amagundane. Kwi-dorsal hairy skin of mouse, iintlobo ezintathu eziphambili zeenwele zichazwe: i-zigzag (malunga ne-72%), i-awl / auchene (malunga ne-23%) kunye nomlindi okanye i-tylotrich (malunga ne-5%).11-14 Zigzag kunye ne-Awl / i-auchenne hair follicles ivelisa i-shafts yeenwele ezincinci kunye nezifutshane kwaye zidibene ne-sebaceous gland enye. Umlindi okanye iinwele ze-tylotrich zezona zinde kwiintlobo zeenwele zeenwele. Zibonakaliswe ngebhalbhu yeenwele ezinkulu ezinxulumene namadlala amabini e-sebaceous. Iinwele zokugada kunye ne-awl / auchene zicwangciswe ngokuphindaphindayo, ipateni ehlala izithuba ngelixa iinwele ezigosozi zihlala kwindawo ezixineneyo zolusu ezijikeleze iindidi ezimbini zeenwele ezinkulu [Fig. 1 (A1, A2 kunye ne-A3)].
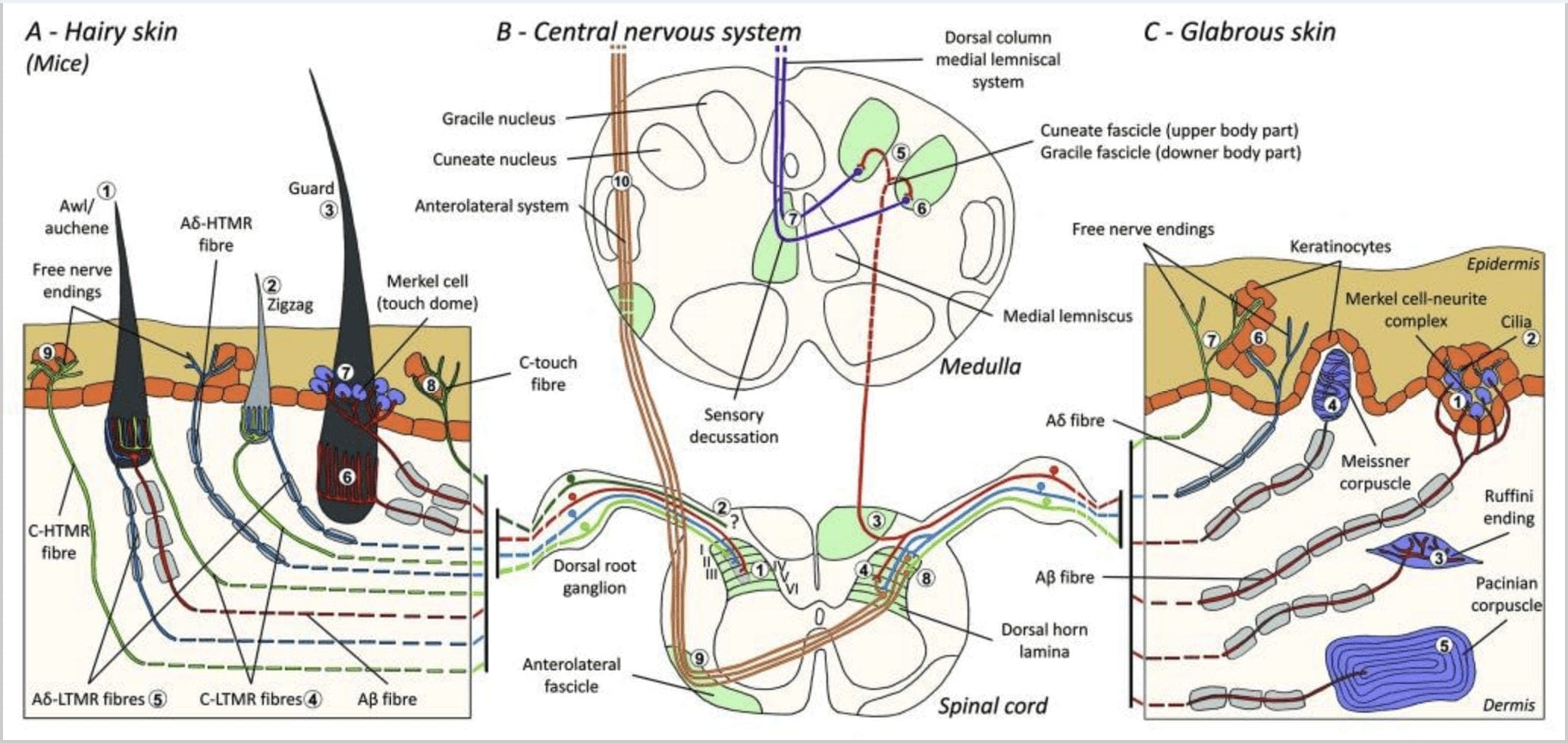
Umzobo 1. Ulungelelwaniso kunye noqikelelo lwe-cutaneous mechanoreceptors. Kwisikhumba esineenwele, i-brush elula kunye ne-touch ibonwa ngokuyinhloko ngokungena ngaphakathi kwi-follicles yeenwele: i-awl / auchenne (A1), i-zigzag (A2) kunye nomlindi (A3). Iinwele zeAwl/auchene ziphindwe kathathu innervated yiC-LTMR lanceolate endings (A4), A?-LTMR and A? ukulungelelanisa ngokukhawuleza-LTMR (A6). Iifollicle zeenwele ze-Zigzag ziinwele zeenwele ezimfutshane kwaye zingenasiphelo kuzo zombini i-C-LTMR (A4) kunye ne-A? -Iziphelo ze-LTMR ze-lanceolate (A5). Ezona follicles zeenwele ezinde zigcinwe ngu-A? ukuziqhelanisa ngokukhawuleza-iziphelo ze-LTMR ze-longitudinal lanceolate (A6) kwaye zihambelana no-A? iqhelanisa ngokucothayo-LTMR yeziphelo zedome yokuchukumisa (A7). Uqikelelo olusembindini wazo zonke ezi ntsinga ziphela ngokwahlukileyo, kodwa ngokuyinxenye i-laminae ewela i-spinal cord dorsal horn (C-LTMR kwi-lamina II, A?-LTMR kwi-lamina III kunye ne-A?-LTMR kwi-lamina IV kunye ne-V). Uqikelelo lwe-LTMR olungenalo i-innervate efanayo okanye i-follicle yeenwele ezikufutshane zilungelelaniswa ukuze zenze ikholamu encinci kwi-spinal cord dorsal horn (B1 ngegrey). Kuphela kwesikhumba esinoboya, i-subpopulation ye-C-fibers isiphelo esikhululekile singenasiphelo kwi-epidermis kwaye siphendule ngokuchukumisa okumnandi (A8). Ezi ntsinga ze-C-touch aziphenduli kwi-touch eyingozi kwaye ukuhamba kwazo kwendlela akukaziwa (B2). Kulusu olukhazimlayo, ukubamba okungenabungozi kulamlwa ziindidi ezine ze-LTMR. I-Merkel cell-neurite complex ikwi-basal layer ye-epidermis (C1). Le mechanoreceptor iquka ilungiselelo phakathi kweeseli ezininzi ze-Merkel kunye ne-terminal ye-nerve eyandisiweyo ukusuka kwi-A enye? ifayibha. Iiseli ze-Merkel zibonisa umnwe njengeenkqubo zokuqhagamshelana ne-keratinocytes (C2). Isiphelo seRuffini sibekwe kwindawo kwidermis. Sisiva esicekethekileyo esimilise okwesiga esifakwe iziphelo zoluvo ezidityaniswe ku-A? ifayibha (C3). I-Meissner corpuscle edityaniswe ku-A? i-nerve end kwaye ifumaneka kwidermal papillae. Le mechanoreceptor ehlanganisiweyo iqukethe iiseli ezixhasayo ezipakishwe phantsi ezilungiselelwe njenge-lamellae ethe tyaba ejikelezwe zizicubu ezixhumeneyo (C4). I-Pacinian corpuscle yi-mechanoreceptor enzulu. Omnye uA? Isiphelo se-nerve esingenamyelini siphela embindini wale corpuscle enkulu ye-ovoid eyenziwe nge-lamellae edibeneyo. Uqikelelo lwezi ntsinga ze-A?-LTMR kwintambo yomqolo zohlulwe ngokwamasebe amabini. I-principal central branch (B3) inyuka kwintambo yomgogodla kwi-ipsilateral dorsal eyenza i-cuneate okanye i-gracile fascicles (B5) kwinqanaba le-medulla apho i-primary afferents yenza i-synapse yabo yokuqala (B6). I-neurons yesibini yenza i-sensory decussation (B7) ukwenza isicatshulwa kwi-lemniscus ephakathi ekhuphuka nge-brainstem ukuya kwi-midbrain, ngokukodwa kwi-thalamus. I-secondary branche ye-LTMR iphelisa uphondo lwe-dorsal kwi-lamina II, IV, V kwaye iphazamise ukuhanjiswa kweentlungu (B4). Ukuchukumisa okukrakra kufunyenwe yi-nerve yamahhala ephela kwi-epidermis yezombini ezinoboya (A9) kunye ne-glabrous skin (C7). Ezi mechanoreceptors zisiphelo se-A?-HTMR kunye ne-C-HTMR ngokusondeleyo kunye neekeratinocytes ezingabamelwane (C6). A?-hTMR iphelisa kwi-lamina I kunye ne-V; I-C-HTMR iphelisa kwi-lamina I kunye ne-II (B8). Kwinqanaba le-spinal cord dorsal horn, ii-primary afferents ze-HTMRs zenza i-synapses kunye ne-neurons yesibini ewela umgca ophakathi kwaye ikhuphukele kwisakhiwo esiphezulu sobuchopho kwi-fascicle anterolateral (B9, B10).
Kungekudala, i-Ginty kunye nabasebenzisana nabo basebenzise ukudibanisa kwe-molecular-genetic labeling kunye ne-somatotopic retrograde tracing approach to viewing the organisation of the peripheral and central axonal endings of LTMRs kwiigundane.15 Iziphumo zabo zixhasa imodeli apho iimpawu zomntu ngamnye ze-complex tactile stimulus are. itsalwa ziintlobo ezintathu zeefollicle zeenwele kwaye yahanjiswa ngemisebenzi yendibaniselwano eyodwa ye-A?-, A?- kunye ne-C-fibers ukuya kwi-dorsal horn.
Baye babonisa ukuba i-genetic labeling ye-tyrosine hydroxylase positive (TH +) i-DRG neurones ibonakalisa i-neuron ye-nonpeptidergic, i-diameter encinci ye-sensory neurones kwaye ivumela ukubonwa kwe-C-LTMR ye-peripheral endings eluswini. Okumangalisa kukuba, amasebe e-axoneal e-C-LTMRs nganye afunyenwe ukuba ahlasele kwaye enze iziphelo ze-longitudinal lanceolate ezihambelana ngokusondeleyo kunye ne-zigzag (i-80% yeziphetho) kunye ne-awl / auchene (i-20% yokuphela), kodwa kungekhona i-tylotrich hair follicles [Fig. 1 (A4)]. Iziphelo ze-Longitudinal lanceolate ziye zacingelwa ukuba zezodwa ze-A?-LTMRs kwaye ngoko ke bekungalindelekanga ukuba iziphelo ze-C-LTMR ziza kwenza i-longitudinal lanceolate endings. i-myelinated mechanoreceptors [Umfanekiso. 15 (C2)].
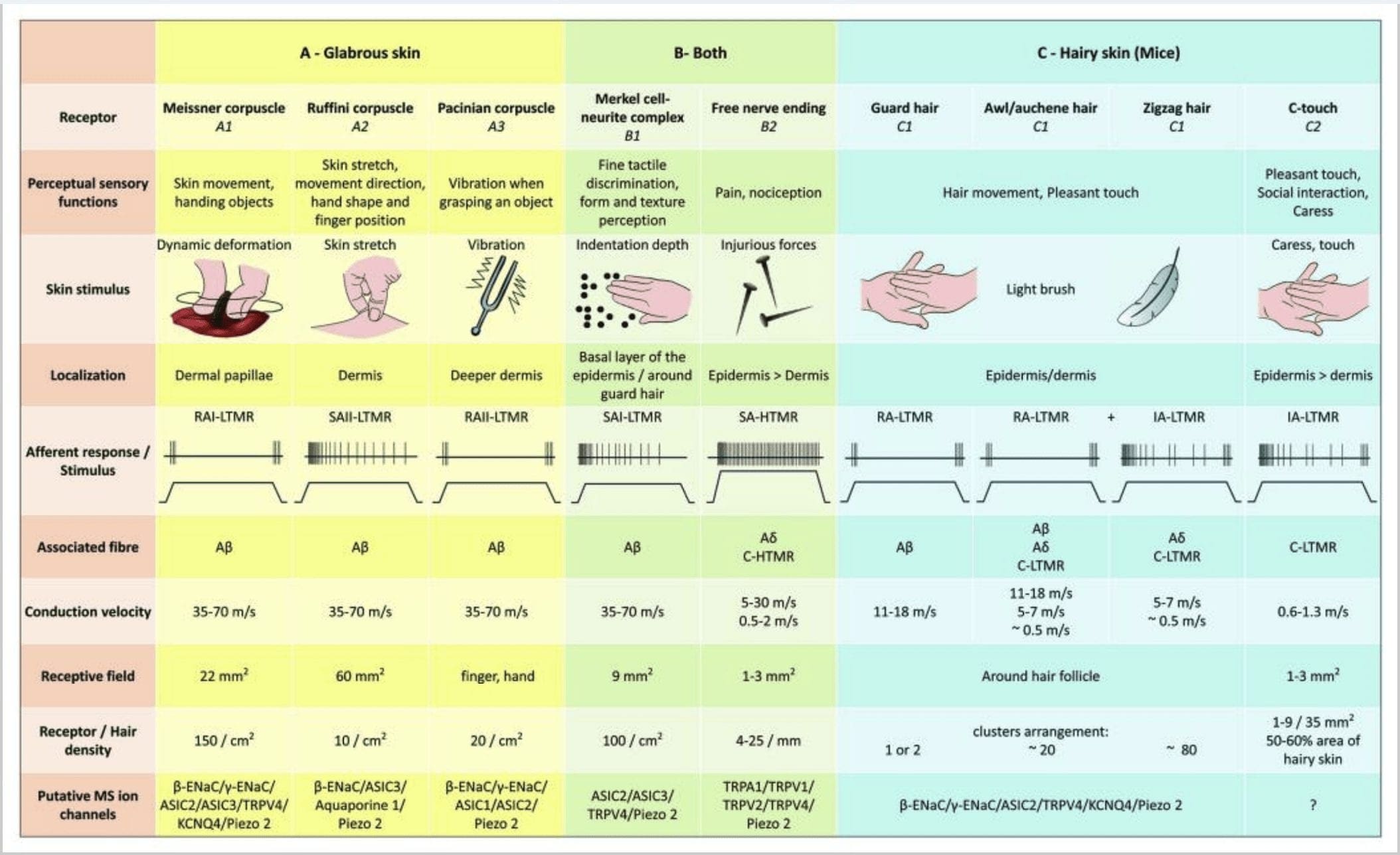
Umzobo 2. I-Tactile receptors kwizilwanyana ezincelisayo: I-tactile receptors ezithintekayo ziyahlula kwi-touch innocuous exhaswa ngama-receptors amaninzi ane-low mechanical threshold (LTMRs) kwi-glabrous and hairy skin and noxious touch exhaswa yi-high mechanical threshold receptor (HTMRs). Benza iziphelo ezingenayo imithambo-luvo eziphela ikakhulu kwi-epidermis. (A) Ulusu olukhazimlayo. I-A1: I-Meissner corpuscles ibona intshukumo yesikhumba kunye nokutyibilika kwento esandleni. Zibalulekile ekunikezeni into kunye ne-dexterity. Ii-receptors zilungelelanisa ngokukhawuleza kwi-stimulus, zixhunywe kwi-A? iifibers kwaye kancinane ukuya kwi-C fibers kwaye ibe nentsimi enkulu ye-receptor. I-A2: I-Ruffini corpuscles ibona ukunweba kwesikhumba kwaye kubalulekile ukufumanisa indawo yeminwe kunye nokunikezela into. I-receptor iqhelana ngokuthe ngcembe kwi-stimulus kunye nomsebenzi ogciniweyo logama nje isivuseleli sisetyenzisiwe. Izamkeli ziqhagamshelwe ku-A? iifibers kwaye zinendawo enkulu yokwamkela. I-A3: I-Pacinian corpuscles inzulu kwi-dermis kwaye ibona ukungcangcazela. Izamkeli ziqhagamshelwe ku-A? imicu; bakhawuleza baziqhelanise novuselelo kwaye babe neyona ntsimi inkulu yolwamkelayo. (B) Ulusu lonke. B1: I-Merkel-cell complexes ikhona kuzo zombini ulusu olukhazimlayo kunye neenwele ezijikeleze. Zibonakaliswe ngokuxineneyo esandleni kwaye zibalulekile ekuboneni ukuthungwa kunye nocalulo olugqwesileyo phakathi kwamanqaku amabini. Banoxanduva lokuchaneka kweminwe. Izamkeli ziqhagamshelwe ku-A? imicu; ngokucothayo baziqhelanisa novuselelo kwaye banentsimi emfutshane eyamkelayo. I-B2: I-HTMRs ye-Noxious touch kunye nokulungelelaniswa okucothayo kakhulu kwi-stimulus, oko kukuthi, esebenzayo nje ngokuba i-nociceptive stimulus isetyenziswa. Zenziwa yi-nerve ephela ye-A? kunye ne-C-fibers ezinxulumene ne-keratinocytes. (C) Ulusu oluneenwele. C1: Iinwele zeenwele zidibene neentlobo ezahlukeneyo zeenwele. Kwiimpuku Iinwele zokugada zezona zinde kwaye zibonakaliswe kancinci, i-awl/auchenne zinobungakanani obuphakathi kwaye i-zigzag zezona zincinci kwaye zezona nwele zixineneyo. Ziqhagamshelwe ku-A? iintsinga kodwa nakwi-A? kunye ne-C-LTMRs iifayili ze-awl / auchenne kunye nezag iinwele. Babona ukunyakaza kweenwele kubandakanya ukubamba okumnandi ngexesha lokuphululwa. Baziqhelanisa ngokukhawuleza okanye nge-intermediate kinetic kwi-stimulus. I-C2: I-C-touch nerve endings ihambelana ne-subtype ye-C fibers terminus enesiphelo samahhala esibonakaliswe ngumgangatho ophantsi womatshini. Bafanele ukuba bafake ikhowudi yemvakalelo emnandi eyenziwa yi-caress. Baziqhelanisa ngokuphakathi kwi-stimulus kwaye banentsimi emfutshane eyamkelayo. Iziteshi ze-ion ze-Pututive mechanosensitive (MS) ezichazwe kwii-receptors ezahlukeneyo ze-tactile ziboniswa ngokufanelekileyo kwiinkcukacha zangaphambili kwaye zishwankathele i-hypothesis ekhoyo phantsi kovavanyo.
Abemi besibini abakhulu abachongiweyo baxhalabisa i-A?-LTMR endings kwi-Awl/Auchenne kunye ne-zigzag follicles ukuba ithelekiswe kunye ne-Down hair follicle efundwe ngokubanzi kwikati kunye nomvundla. I-Ginty kunye nabadibanisi babonise ukuba i-TrkB ibonakaliswe kumanqanaba aphezulu kwi-subset ye-DRG neurones ephakathi. Ukurekhodwa kwe-intracellular kusetyenziswa i-ex vivo skin-nerve yokulungiselela iifiber ezibhaliweyo zibonise ukuba zibonisa iimpawu ze-physiological of fibers ezifundwe ngaphambili kwikati kunye nomvundla: i-exquisite mechanical sensitivity (i-Von Frey threshold <0.07 mN), ngokukhawuleza ukulungelelanisa iimpendulo kwi-suprathreshold stimuli, ukuqhuba okuphakathi. isantya (5.8 � 0.9 m/s) kunye ne-soma spikes emxinwa engashukumiyo.15 Ezi A?-LTMRs zenza iziphelo ze-longitudinal lanceolate ezinxulumene phantse nayo yonke i-zigzag kunye ne-awl/auchene follicle yeenwele zesiqu [Fig. 1 (A5)].
Ekugqibeleni, babonisa ukuba iziphelo zepheripheral zokuziqhelanisa ngokukhawuleza no-A? I-LTMRs yenza iziphelo ze-longitudinal lanceolate ezinxulumene nomlindi (okanye i-tylotrich) kunye ne-awl / auchene iinwele zeenwele [Umfanekiso. 1 (A6)].15 Ukongeza, iinwele zomlindi zikwanxulunyaniswa nekhompleksi yeseli ye-Merkel eyenza idome yokucofa eqhagamshelwe ku-A? ukulungelelanisa ngokucothayo i-LTMR [Fig. 1 (A7)].
Isishwankathelo, phantse zonke iifollicle zeenwele ze-zigzag zi-innervated yi-C-LTMR kunye ne-A?-LTMR iziphelo ze-lanceolate; Iinwele/iinwele ze-auchene zibanjwa kathathu ngu-A? ukulungelelanisa ngokukhawuleza-LTMR, A?-LTMR kunye neC-LTMR isiphelo se-lanceolate; Iifollicle zeenwele zokugada zigcinwe ngu-A? ukuziqhelanisa ngokukhawuleza-LTMR longitudinal lanceolate iziphelo kunye nokusebenzisana no-A? iqhelanisa ngokucothayo-LTMR yeziphelo zedome yokuchukumisa. Ngaloo ndlela, i-follicle nganye yeenwele zemouse ifumana indibaniselwano ekhethekileyo kunye nezingaguqukiyo ze-LTMR eziphela ezihambelana ne-neurophysiologically distinct mechanosensory end parts. Ukuthathela ingqalelo ukulungiswa okuphindaphindiweyo kwezi ntlobo zintathu zeenwele, u-Ginty kunye nabadibanisi bacebisa ukuba ulusu olunoboya lubandakanya ukuphindaphinda okuphindaphindiweyo kweyunithi yombhobho equlethe, (1) enye okanye ezimbini iinwele zonogada ezibekwe kwindawo esembindini, (2) ~20 ezingqonge i-awl/auchenne iinwele kunye (3) ) Iinwele ezingama-80 ezidibeneyo ezigoso-goso [Fig. 2 (C1)].
Intelekelelo yentambo yomqolo. Uqikelelo olusembindini we-A? ukulungelelanisa ngokukhawuleza-LTMRs, A?-LTMRs kunye ne-C-LTMRs ziphela ngokwahlukileyo, kodwa zigqithise i-laminae (II, III, IV) ye-spinal cord dorsal horn. Ukongezelela, ii-terminals eziphambili ze-LTMRs ezingenayo i-innervate efanayo okanye i-follicle yeenwele ezikufutshane ngaphakathi kweyunithi ye-LTMR ye-peripheral ihambelana nokwenza ikholamu ye-LTMR encinci kwi-spinal cord dorsal horn [Umfanekiso. 1 (B1)]. Ngaloo ndlela, kubonakala ngathi i-wedge, okanye ikholamu ye-somatotopically i-primary afferent endings in the dorsal horn imele ukulungelelaniswa kweengqikelelo eziphambili ze-A?-, A? izivuseleli ezisebenza kwiqela elincinane elifanayo leefollicle zeenwele. Ngokusekwe kumanani omlindi, i-awl/ i-auchene kunye neenwele ezizigzag zesiqu kunye namalungu kunye namanani ohlobo ngalunye lwe-LTMR oluncinci, i-Ginty kunye nabasebenzisana nabo baqikelela ukuba uphondo lomqolo wempuku luqulathe i-2,000-4,000 LTMR ikholamu, ehambelana nenani eliqikelelweyo le-peripheral. iiyunithi zeLTMR.15
Ngapha koko, ii-axones ze-LTMR subtypes zinxulunyaniswa ngokusondeleyo, zineengqikelelo ezidibeneyo kunye neziphelo ezidibeneyo ze-lanceolate ezingafaki i-follicle yeenwele ezifanayo. Ukongeza, ngenxa yokuba ezi ntlobo zintathu zeenwele zibonisa iimilo ezahlukeneyo, ubungakanani kunye nokwakheka kweeselfowuni, kunokwenzeka ukuba zibe neempawu eziguqukayo okanye zokungcangcazela. Ezi ziphumo ziyahambelana nemilinganiselo ye-neurophysiological yakudala kwikati kunye nomvundla ebonisa ukuba u-A? I-RA-LTMRs kunye ne-A?-LTMR inokuthi isebenze ngokwahlukileyo ngokuphambuka kweentlobo zeenwele ezihlukeneyo.16,17
Ukuqukumbela, ukubamba kwesikhumba esineenwele kukudibanisa: (1) amanani ahambelanayo, ulwabiwo olulodwa lwesithuba kunye neempawu ezahlukileyo ze-morphological kunye ne-deflectional yeentlobo ezintathu zeefollicle zeenwele; (2) imidibaniso ekhethekileyo ye-LTMR yohlobo oluncinci lweziphelo ezinxulumene nolunye uhlobo lweenwele ezintathu; kunye (3) uvakalelo olwahlukileyo, isantya sokuqhuba, iipateni zikaloliwe we-spike kunye neepropathi zohlengahlengiso kwiiklasi ezine eziphambili ze-LTMRs ezinxulumene neenwele ezenza ukuba inkqubo ye-mechanosensory yesikhumba enoboya ikhuphe kwaye idlulisele kwi-CNS indibaniselwano entsonkothileyo yeempawu ezichaza a touch.
Iziphelo ze-Free-Nerve LTMRs
Ngokubanzi, ii-C-fibers iziphelo zasimahla elulusu zii-HTMRs, kodwa i-subpopulation ye-C-fibers ayiphenduli kuchuku oluyingozi. Le subset ye-tactile C-fiber (CT) afferents imele uhlobo oluhlukileyo lwe-unmyelinated, i-low-threshold mechanoreceptive units ekhoyo kwi-hay hairy kodwa i-glabrous skin yabantu kunye nezilwanyana ezincelisayo [Umfanekiso. I-1 (A8)]. I-18,19 CTs ngokuqhelekileyo idibene nombono wokuvuselela okumnandi kwi-tactile kuqhagamshelwano lomzimba.20,21
I-CT afferents isabela kwi-indentation forces kuluhlu lwe-0.3�2.5 mN kwaye ke ngoko ivakalelwa kakhulu kwi-skin deformation njengoko ininzi ye-A? I-afferents.19 Iimpawu zokulungelelaniswa kwe-CT afferents ngoko ke ziphakathi xa kuthelekiswa ne-mechanoreceptors ye-myelinated ngokukhawuleza nangokukhawuleza. Imimandla eyamkelekileyo ye-CT afferents irhangqe ngokurhangqa okanye i-oval ngokwemilo. Ibala liquka indawo enye ukuya kwezilithoba ezincinci ezisabelayo ezisasazwa kwindawo ukuya kuthi ga kwi-35 mm2.22 Izamkeli ze-homolog zemouse zicwangciswe ngokwepateni yamabala ayekayo agquma malunga ne-50�60% yendawo kwisikhumba esinoboya [Fig. 2 (C2)].23
Ubungqina obuvela kwizigulane ezingenazo i-myelinated tactile afferents zibonisa ukuba ukubonakaliswa kwi-CT fibers kusebenze i-cortex ye-insular. Ekubeni le nkqubo ihlwempuzekile kwi-encoding imiba ecalucalulo yokuchukumisa, kodwa ifanelekile kwi-encoding ecothayo, i-touch touch, i-CT fibers kwisikhumba esinoboya inokuba yinxalenye yenkqubo yokucubungula izinto ezithandekayo kunye nezentlalo ezifanelekileyo zokuthintela. banendima ekuvimbeleni intlungu kwaye kutshanje kucetywayo ukuba ukuvuvukala okanye ukuxhwaleka kunokutshintsha imvakalelo ehanjiswa yi-C-fiber LTMRs ukusuka ekuthinteni okumnandi ukuya kwintlungu.24
Yeyiphi indlela yokuhamba nge-CT-afferents engekaziwa [Fig. I-1 (B2)], kodwa i-low-threshold tactile igalelo kwiiseli ze-spinothalamic projection ziye zabhalwa, i-27 yokuboleka ukunyaniseka kwiingxelo ezifihlakeleyo, eziphikisanayo zokufunyanwa kwe-touch kwizigulane zabantu emva kokutshatyalaliswa kwezi ndlela emva kweenkqubo ze-chordotomy.28
LTMRs kwi-Glabrous Skin
I-Merkel cell-neurite complexes kunye ne-dome yokuchukumisa. UMerkel (1875) waba ngowokuqala ukunika inkcazo yembali yeeseli ze-epidermal ezine-nuclei ezinkulu eziqhagamshelweyo, enxibelelana ne-afferent nerve fibers ecingelwayo. Wayecinga ukuba bagcina imvakalelo yokuchukumisa ngokubabiza ngokuba yiTastzellen (iiseli ezibambekayo). Ebantwini, ii-cell ze-Merkel ze-neurite complexes zityetyiswe kwiindawo ezithintekayo zolusu, zifumaneka kwi-basal layer ye-epidermis kwiminwe, imilebe kunye nezitho zangasese. Zikwakho kwisikhumba esinoboya kubuninzi obusezantsi. I-Merkel cell-neurite complex iquka iseli ye-Merkel ekufutshane ne-terminal yemithambo-luvo esuka kwi-myelinated A enye? ifayibha [Fig. I-1 (C1)] (uphononongo kwi-Halata kunye nabasebenzisana nabo) .29 Kwicala le-epidermal iseli ye-Merkel ibonisa iinkqubo ezinjengomnwe ezidlula phakathi kwee-keratinocyte ezingabamelwane [Fig. 1 (C2)]. Iiseli ze-Merkel ziyi-keratinocyte-derived epidermal cells.30,31 Ixesha le-touch dome laqaliswa ukubiza i-concentration enkulu ye-Merkel cell complexes kwisikhumba esinoboya se-cat forepaw. I-touch dome inokuba ne-150 yeeseli ze-Merkel ezingavalelwanga yi-A?-fiber enye nasebantwini ngaphandle kwe-A?-fibers, A? kunye neeC-fibers zazikhona rhoqo.32-34
Uvuselelo lwe-Merkel cell�neurite complexes lukhokelela kwiimpendulo zoHlobo loku-I (SA I) oluziqhelanisa kancinci kancinci, olusuka kwiindawo zolwamkelo zeziphumlisi ezinemida ebukhali. Akukho kukhutshwa okuzenzekelayo. Ezi zinkimbinkimbi ziphendula kubunzulu be-indentation kwesikhumba kwaye zinesisombululo esiphezulu sendawo (0.5 mm) ye-cutaneous mechanoreceptors. Basasaza umfanekiso ochanekileyo wendawo ye-tactile stimuli kwaye bacetywa ukuba babe noxanduva lokumila kunye nokuthungwa kocalulo [Fig. 2 (B1)]. Iimpuku ezingenazo iiseli zeMerkel azikwazi ukubona umphezulu owenziwe ngeenyawo ngeenyawo ngelixa zisenza oko zisebenzisa amabhovu azo.35
Ingaba iseli ye-Merkel, i-neuron ye-sensory okanye zombini iindawo ze-mechanotransduction isengumcimbi wengxoxo. Kwiigundane, ukutshatyalaliswa kwe-phototoxic yeeseli ze-Merkel kutshitshisa impendulo ye-SA I.36 Kwiimpuku ezineeseli ze-Merkel ezicinezelwe ngokwemfuza, impendulo ye-SA I erekhodwe kwi-ex vivo yesikhumba / ukulungiswa kwenzwa yanyamalala ngokupheleleyo, ebonisa ukuba iiseli ze-Merkel ziyafuneka kwi-encoding efanelekileyo ye-Merkel. iimpendulo ze-receptor.37 Nangona kunjalo, ukuvuselela umatshini weeseli ze-Merkel ezizimeleyo kwinkcubeko ngoxinzelelo oluqhutywa yimoto aluvelisi i-currents mechanical-gated currents.38,39 Keratinocytes inokudlala indima ebalulekileyo ekusebenzeni okuqhelekileyo kwe-cell Merkel cell�neurite complex. Iinkqubo ze-Merkel ezinjengomnwe womnwe zingahamba kunye ne-skin deformation kunye ne-epidermis cell movement, kwaye le nto ingaba yinyathelo lokuqala lokutshintshwa komatshini. Ngokucacileyo, iimeko ezifunekayo ukufunda i-mechano-sensitivity yeeseli ze-Merkel azikasekwa.
Iziphelo zeRuffini. Iziphelo zeRuffini ziziphelo zoluvo ezicekethekileyo ezimilise okwesiga ezidityaniswe ku-A? iziphelo zemithambo-luvo. Iziphelo zeRuffini zizisilinda zeethishu ezincinci ezidityanisiweyo ezicwangciswe kunye nedermal collagen strands ezibonelelwa nge-myelinated nerve fibers enye ukuya kwezintathu ze-4�6 �m diametre. Ukuya kuthi ga kwiisilinda ezintathu zokuqhelaniswa okwahlukileyo kwidermis zinokudityaniswa zenze isamkeli esinye [Umfanekiso. 1 (C3)]. Ngokwesakhiwo, iziphelo zeRuffini zifana nezitho ze-tendon zeGolgi. Zibonakaliswa ngokubanzi kwidermis kwaye zichongiwe njengohlobo lwe-II oluthambileyo (SA II) lwe-cutaneous mechanoreceptors. Ngokuchasene nemvelaphi yomsebenzi we-nervous ngokuzenzekelayo, ukukhutshwa okuthe chu-ukuqhelanisa rhoqo kubangelwa yi-perpendicular low force egcinwe ukuvuselela ngoomatshini okanye ngokusebenzayo ngakumbi nge-dermal stretch. Impendulo ye-SA II isuka kwiindawo ezinkulu zolwamkelo ezinemida engacacanga. I-Ruffini receptors inegalelo ekubonweni kwesalathiso sento yokuhamba ngepatheni yokwelula kwesikhumba [Umfanekiso. 2 (A2)].
Kwiigundane, iimpendulo ze-SA I kunye ne-SA II zinokuhlukaniswa nge-electrophysiologically kwi-ex-vivo nerve-skin preparation. ukugcinwa kwebhalansi ye-osmotic yamazinyo eyimfuneko kwi-mechanoTransduction.
Meissner iicorpuscles. I-Meissner corpuscles ibekwe kwindawo kwi-dermal papillae yolusu olukhazimlayo, ikakhulu kwiintende zezandla kunye neenyawo kodwa nakwimilebe, ngolwimi, ebusweni, kwiingono nakwizitho zangasese. Ngokwe-anatomically, zibandakanya i-nerve encapsulated ending, i-capsule yenziwe ngeeseli ezixhasayo ezicwangcisiweyo ezicwangciswe njenge-lamellae ethe tye efakwe kwiithishu ezidibeneyo. Ngaba kukho i-fiber ye-nerve enye A? ii-afferents eziqhagamshelwe kwi-corpuscle nganye [Fig. 1 (C4)]. Nakuphi na ukuguqulwa komzimba we-corpuscle kubangela i-volley yezenzo ezinokuthi ziphele ngokukhawuleza, oko kukuthi, zilungelelanisa ngokukhawuleza ii-receptors. Xa i-stimulus isusiwe, i-corpuscle iphinda ibuyele kwimo yayo kwaye ngelixa isenza njalo ivelisa enye i-volley yezenzo ezinokwenzeka. Ngenxa yendawo engaphezulu kwidermis, ezi corpuscles zisabela ngokukhethayo kwintshukumo yesikhumba, ukubonwa okubambekayo kokutyibilika kunye nokungcangcazela (20�40 Hz). Zinobuntununtunu kulusu oluguquguqukayo - umzekelo, phakathi kwesikhumba kunye nento ephathwayo [Fig. 2 (A1)].
I-Pacinian corpuscles. I-Pacinian corpuscles ziyi-mechanoreceptors ezinzulu zolusu kwaye zezona zithintekayo ze-cutaneous mechanoreceptor ye-skin motion. Ezi ngqungquthela ezinkulu ze-ovoid (i-1 mm ubude) zenziwe nge-lamellae edibeneyo ye-fibrous connective tissue kunye ne-fibroblasts ehlanganiswe ngamaseli e-Schwann aguquliweyo abonakaliswe kwi-deep dermis. , iphelisa u-A omnye? i-afferent unmyelinated nerve ephela [Fig. 43 (C1)]. Zinebala elikhulu lolwamkelo kumphezulu wolusu oneziko elinovakalelo ngokukodwa. Uphuhliso kunye nomsebenzi weentlobo ezininzi ze-mechanoreceptor ezilungelelanisa ngokukhawuleza ziphazamiseka kwiigundane ze-c-Maf eziguqukayo. Ngokukodwa, i-Pacinian corpuscles i-atrophied kakhulu.5
I-Pacinian corpuscles ibonisa ukulungelelaniswa ngokukhawuleza kakhulu ekuphenduleni ukunyanzeliswa kwesikhumba, i-II (RA II) yokukhupha ngokukhawuleza i-nervous discharge ekwazi ukulandela i-frequency ephezulu ye-vibratory stimuli, kwaye ivumele ukubonwa kweziganeko ezikude ngokugungqa.45 I-Pacinian corpuscle. i-afferents iphendula kwi-indentation eqhubekayo kunye nomsebenzi wexeshana ekuqaleni kunye nokunciphisa i-stimulus. Zikwabizwa ngokuba yi-acceleration detectors kuba ziyakwazi ukubona utshintsho kumandla okuvuselela kwaye, ukuba izinga lokutshintsha kwi-stimulus liguqulwa (njengoko kwenzeka kwii-vibrations), impendulo yabo ihambelana nolu tshintsho. Ii-Pacinian corpuscles ziva utshintsho olukhulu loxinzelelo kunye noninzi lwazo zonke ii-vibrations (150�300 Hz), ezinokuthi zibhaqe neesentimitha kude [Fig. 2 (A3)].
Impendulo ye-Tonic yabonwa kwi-decapsulated Pacinian corpuscle.46 Ukongezelela, i-Pacinian corpuscles engaguqukiyo iphendula ngomsebenzi oqhubekayo ngexesha le-indentation stimuli, ngaphandle kokuguqula imibundu yomatshini okanye i-frequency yokuphendula xa i-GABA-mediated signaling ivaliwe phakathi kwe-lamellate glia kunye ne-nerve ending.47 Ngaloo ndlela, amacandelo angenayo i-neuronal ye-Pacinian corpuscle inokuba neendima ezimbini ekuhluzeni i-stimulus yomatshini kunye nokulungelelanisa iimpawu zokuphendula ze-neuron ye-sensory.
Uqikelelo lwentambo yomqolo. Uqikelelo lwe-A?-LTMRs kwintambo yomqolo zohlulwe ngokwamasebe amabini. I-principal central branch ikhuphuka kwintambo yomgogodla kwi-ipsilateral dorsal columns ukuya kwinqanaba lomlomo wesibeleko [Umfanekiso. 1 (B3)]. Amasebe asesekondari aphelisa uphondo lwe-dorsal kwi-laminae IV kwaye aphazamise ukuhanjiswa kweentlungu, umzekelo. Oku kunokuthomalalisa iintlungu njengenxalenye yolawulo lwesango [Fig. 1 (B4)].48
Kwimigangatho yomlomo wesibeleko, i-axones yesebe eliyintloko ihluke kwiiphecana ezimbini: i-middleline tract iquka i-fascicle ye-gracile ehambisa ulwazi olusuka kwisiqingatha esisezantsi somzimba (imilenze kunye ne-trunk), kunye necandelo langaphandle liquka i-fascicle ye-cuneate ehambisa ulwazi oluvela kwisiqingatha esiphezulu. yomzimba (iingalo kunye nesiqu) [Fig. 1 (B5)].
I-primary tactile afferents zenza i-synapse yazo yokuqala kunye ne-neurones ye-second order kwi-medulla apho iifayili ezivela kwiphecana nganye ye-synapse kwi-nucleus yegama elifanayo: i-gracile fasciculus axones i-synapse kwi-nucleus ye-gracile kunye ne-cuneate axones synapse kwi-nucleus ye-cuneate [Umfanekiso. 1 (B6)]. I-Neurones efumana i-synapse inikezela nge-afferents yesibini kwaye iwele umgca ophakathi ngokukhawuleza ukwenza iphecana kwicala le-contralateral le-brainstem yengqondo-i-lemniscus ephakathi-enyuka nge-brainstem ukuya kwisikhululo esilandelayo sokuhambisa kwi-midbrain, ngokukodwa, kwi-thalamus [Fig. . 1 (B7)].
Ukucaciswa kwemolekyuli ye-LTMRs. Iinkqubo zeemolekyuli ezilawula ukwahluka kwangaphambili kwe-LTMRs zisandula ukucaciswa ngokuyinxenye. I-Bourane kunye nabasebenzisana nabo baye babonisa ukuba i-neuronal populations evakalisa i-Ret tyrosine kinase receptor (Ret) kunye ne-co-receptor yayo ye-GFR?2 kwi-E11�13 iigundane ze-embryonic DRG ngokukhetha i-coexpress i-transcription factor Mafa.49,50 Aba babhali babonisa ukuba i-Mafa / I-Ret/GFR? I-2 neurones ezimiselwe ukuba zibe ziintlobo ezintathu ezikhethekileyo ze-LTRM ekuzalweni: i-SA1 neurones i-innervating Merkel-cell complexes, i-neurones eguqukayo ngokukhawuleza i-innervating corpuscles ye-Meissner kunye ne-afferents eguqukayo ngokukhawuleza (i-RA I) eyenza isiphelo se-lanceolate malunga neefollicles zeenwele. I-Ginty kunye nabasebenzisana nabo baxela ukuba i-DRG neurones evakalisa ngokukhawuleza i-Ret ilungisa ngokukhawuleza i-mechanoreceptors esuka kwi-Meissner corpuscles, i-Pacinian corpuscles kunye neziphetho ze-lanceolate ezijikeleze iifollicles zeenwele. Uqikelelo lwe-neurone axonal ngaphakathi kwi-brainstem.
Ukuphononongwa kwe-mechanoreceptors yesikhumba somntu. Ubuchwephesha be-�microneurography� echazwe nguHagbarth noVallbo kwi-1968 isetyenziselwe ukufunda ukukhutshwa kokuziphatha kwabantu ababodwa be-mechanosensitive endings ukubonelela izihlunu, ukudibanisa kunye nesikhumba (jonga ukuhlaziywa kweMacefield, 2005) .52,53 Uninzi lwe-microneurography yesikhumba somntu. Izifundo zibonakalise i-physiology ye-tactile afferents kwi-glabrous skin yesandla. Ukurekhodwa kwe-Microelectrode ukusuka kwi-median kunye ne-ulnar nerves kwizifundo zabantu ziye zabonisa i-sensation ye-touch eveliswa ziiklasi ezine ze-LTMRs: I-Meissner afferents ibaluleke kakhulu ekukhanyeni ukukhanya kwesikhumba, ukuphendula kwimikhosi ye-shear yendawo kunye ne-incipient okanye i-overt slips ngaphakathi kwendawo eyamkelayo. Ii-Pacinian afferents zinovelwano ngokugqithisileyo kwi-brisk mechanical transients. I-Afferents iphendula ngamandla ngokuvuthela phezu kwentsimi eyamkelekileyo. I-Pacinian corpuscle ehlala kwidijithi iya kuphendula ngokucofa itafile exhasa ingalo. I-Merkel afferents ineempawu eziphezulu zovakalelo oluguquguqukayo kwi-indentation stimuli esetyenziswa kwindawo ecacileyo kwaye ihlala iphendula ngokukhupha ngaphandle ngexesha lokukhululwa. Nangona i-Ruffini afferents isabela kumandla asetyenziswa ngokuqhelekileyo eluswini, uphawu olulodwa lwe-SA II afferents kukukwazi ukusabela nakwi-lateral skin stretch. Ekugqibeleni, iiyunithi zeenwele kwi-forearm zine-ovoid enkulu okanye imimandla engavumelekanga eyamkelekileyo eyenziwe ngamabala amaninzi abuhlungu ahambelana neenwele zomntu ngamnye (unikezelo ngalunye oluhambelanayo ~ 20 iinwele).
Ukuziva kweeMechanical zeKeratinocytes
Naliphi na i-mechanical stimulus eluswini kufuneka ihanjiswe ngeekeratinocytes ezenza i-epidermis. Ezi seli zifumaneka kuyo yonke indawo zinokwenza imisebenzi yokubonisa ukongeza kwiindima zabo ezixhasayo okanye ezikhuselayo. Ngokomzekelo, i-keratinocytes ikhupha i-ATP, i-molecule ebalulekileyo ye-sensory signaling, ekuphenduleni i-mechanical and osmotic stimuli.54,55 Ukukhululwa kwe-ATP kubangela ukwanda kwe-calcium ye-intracellular ngokuvuselela i-autocrine ye-purinergic receptors.55 Ngaphezu koko, kukho ubungqina bokuba i-hypotonicity ivuselela i-Rho. -i-kinase indlela yomqondiso kunye ne-F-actin elandelayo yoxinzelelo lwe-fiber formation ebonisa ukuba ukuguqulwa komatshini we-keratinocytes kunokuphazamisa ngomatshini kunye neeseli zommelwane ezifana neeseli ze-Merkel zokuchukumisa ezingenabungozi kunye ne-C-fiber eziphelweni zamahhala zokuthintela okubi [Umfanekiso. 1 (C6)].56,57
Ukuchukumisa okuNoxious
I-high threshold mechanoreceptors (HTMRs) yi-epidermal C- kunye ne-A? free nerve-iziphelo. Azihambelani nezakhiwo ezikhethekileyo kwaye zibonwa kwisikhumba esineenwele zombini [Umfanekiso. 1 (A9)] kunye nolusu olukhazimlayo [Umfanekiso. 1(C7)]. Nangona kunjalo, ixesha lokuphela kwemithambo-luvo kufuneka lithathelwe ingqalelo ngobulumko kuba iziphelo zemithambo-luvo zihlala zisondelelene ne-keratinocyte okanye iseli yeLangherans okanye i-melanocytes. Uhlalutyo lwe-Ultrastructural lwe-nerve endings lubonisa ubukho be-endoplasmic reticulum, i-mitochondria eninzi kunye ne-dense-core vesicle. Iimbumba ezikufutshane zeeseli ze-epidermal ziqina kwaye zifana ne-post-synaptic membrane kwizicubu ze-nervous. Qaphela ukuba ukusebenzisana phakathi kwe-nerve endings kunye neeseli ze-epidermal zinokuba zimbini kuba iiseli ze-epidermal zinokukhulula abalamli njenge-ATP, i-interleukine (IL6, i-IL10) kunye ne-bradykinin kunye ne-peptidergic nerve endings inokukhulula iipeptides ezifana ne-CGRP okanye i-substance P esebenza kwiiseli ze-epidermal. I-HTMRs iquka i-mechano-nociceptors evuyiswa kuphela yi-noxious mechanical stimuli kunye ne-polymodal nociceptors eziphinda ziphendule ukushisa okukrakra kunye neekhemikhali zangaphandle [Umfanekiso. 2 (B2)].58
I-HTMR i-afferent fibers iphelisa kwi-projection neurones kwi-dorsal horn ye-spinal cord. I-A?-HTMRs iqhagamshelana ne-second order neurones kakhulu kwi-lamina I kunye ne-V, kanti i-C-HTMRs iphela kwi-lamina II [Fig. 1 (B8)]. Iprojekthi yesibini ye-nociceptive neurones ukuya kwicala lokulawula intambo yomgogodla kwaye inyuke kumbandela omhlophe, yenze inkqubo ye-anterolateral. Ezi neurones ziphela ikakhulu kwithalamus [Fig. 1 (B9 kunye ne-B10)].
I-Mechano-Currents kwi-Somatosensory Neurones
Iindlela zokucotha okanye ukulungelelaniswa ngokukhawuleza kwe-mechanoreceptors azikacaciswa. Akucaci ukuba ingakanani i-mechanoreceptor adaptation inikezelwa yimeko yeselula ye-nerve ye-sensory ending, i-intrinsic properties of the mechanical-gated channels and properties of the axonal voltage-gated ion channels in sensory neurones (Fig. 2). Nangona kunjalo, inkqubela phambili yamva nje ekubonakalisweni kwee-currents-gated currents ibonise ukuba iiklasi ezahlukeneyo ze-mechanosensitive channels zikhona kwi-DRG neurones kwaye zingachaza ezinye iinkalo zokulungelelanisa i-mechanoreceptors.
Ukurekhoda kwe-in vitro kwiigundane kubonise ukuba i-soma ye-DRG neurons i-intrinsically mechanosensitive kwaye ivakalisa i-cationic mechano-gated currents. kunye ne-benzamil, kubangela i-block block.59 FM64-60,62,63 isebenza njenge-blocker ehlala ixesha elide, kunye ne-injection ye-FM1-43 kwi-find paw yeempuku kunciphisa uvakalelo lwentlungu kuvavanyo lwe-Randall-Selitto kunye nokwandisa i-paw threshold ehlolwe nge von Frey iinwele.1
Ekuphenduleni ukuvuselelwa komatshini, i-mechanosensitive currents iyancipha ngokuvalwa. Ngokusekelwe kwixesha eliqhubekayo lokubola kwangoku, iintlobo ezine ezihlukeneyo zemisinga ye-mechanosensitive ziye zahlula: ukulungelelanisa ngokukhawuleza imisinga (~3�6 ms), ukulungelelaniswa kwemisinga (~15�30 ms), ukulungelelanisa ngokukhawuleza imisinga (~200�300 ms). ) kunye ne-ultra-slowly adapting currents (~ 1000 ms) .64 Yonke le misinga ikhona kunye neziganeko eziguquguqukayo kwi-rat DRG neurones innervating skin glabrous of the hindpaw.64
Uvakalelo lomatshini lwemisinga ye-mechanosensitive lunokumiselwa ngokusebenzisa uchungechunge lwe-stimuli eyongeziweyo yomatshini, ukuvumela uhlalutyo olucacileyo lwe-stimulus-yangoku.66 Ulwalamano lwangoku oluvuselelayo luqheleke njenge-sigmoidal, kwaye ubuninzi be-amplitude yangoku bunqunywe linani iziteshi ezivulekileyo ngaxeshanye.64,67 Okubangel 'umdla kukuba, ukulungelelaniswa kwe-mechanosensitive yangoku kuye kwabikwa ukuba kubonise umda ophantsi we-mechanical threshold kunye ne-half-activation midpoint xa kuthelekiswa ne-ultra-slowly adapting mechanosensitive current.63,65
I-neurones ye-Sensory kunye ne-non-nociceptive phenotypes ekhethwayo ivakalisa ngokukhawuleza ukulungelelanisa i-mechanosensitive currents kunye ne-low threshold mechanical threshold.60,61,63,64,68 Ngokuchaseneyo, ngokukhawuleza kunye ne-ultra-slowly adapting mechanosensitive currents zichazwe ngamaxesha athile kwiiseli ze-putive non-nociceptive.64,68 Oku kubangele isiphakamiso sokuba le misinga ingaba negalelo kwimigangatho eyahlukeneyo yomatshini ebonwa kwi-LTMRs kunye ne-HTMRs kwi-vivo. Nangona ezi zilingo ze-in vitro kufuneka zithathwe ngokuqaphela, ukuxhaswa kobukho kwi-soma ye-DRG neurones ye-mechanotransducers ephantsi kunye ne-high-threshold ye-mechanotransducers nayo ibonelelwe nge-radial stretch-based stimulation of cultured mouse sensory neurones.69 Le paradigm ibonakalise ezimbini. abantu abaphambili bee-neurones ezizolulayo, enye esabela kwi-low stimulus amplitude kunye nenye ephendula ngokukhethayo kwi-amplitude ephezulu yokuvuselela.
Ezi ziphumo zineempembelelo ezibalulekileyo, kodwa eziqikelelwayo, ze-mechanistic: i-mechanical threshold ye-sensory neurones inokuthi ingabinanto encinci yokwenza ne-cell organisation ye-mechanoreceptor kodwa inokuthi ilale kwiipropati ze-ion channels mechanical-gated.
Iinkqubo ezenza i-desensitization ye-mechanosensitive cation currents kwi-rat DRG neurones zisandul 'ukutshatyalaliswa.64,67 Iphuma kwiindlela ezimbini ezihambelanayo ezichaphazela iipropati zesiteshi: ukulungelelaniswa kunye nokungasebenzi. Ukuziqhelanisa kwaxelwa okokuqala kwizifundo zeeseli zeenwele. Inokuchazwa ngokusebenza njengenguqulelo elula ye-transducer channel's activation curve ecaleni kwe-mechanical stimulus axis.70-72 Ukulungelelaniswa kuvumela i-receptors ye-sensory ukuba igcine uvakalelo lwabo kwi-stimuli entsha phambi kwe-stimulus ekhoyo. Nangona kunjalo, iqhezu elikhulu lemisinga ye-mechanosensitive kwi-DRG neurones ayinakuphinda isebenze ngokulandela ukuvuselela ukuguqulwa komatshini, okubonisa ukungasebenzi kwezinye iziteshi ze-transducer.64,67 Ngoko ke, zombini ukungasebenzi kunye nokulungelelanisa isenzo kwi-tandem yokulawula imisinga ye-mechanosensitive. Ezi ndlela zimbini ziqhelekileyo kuzo zonke ii-currents ze-mechanosensitive ezichongiweyo kwi-rat DRG neurones, ebonisa ukuba izinto ezinxulumene ne-physicochemical zinquma i-kinetics yale mijelo.64
Ukuqukumbela, ukumisela iipropati ze-endogenous mechanosensitive currents in vitro kubalulekile kumzamo wokuchonga iindlela zokutshintshwa kwinqanaba le-molecular. Ukwahlukahlukana okubonwe kwi-threshold yomatshini kunye nokulungelelanisa i-kinetics yeendlela ezahlukeneyo ze-mechanical-gated currents kwi-DRG neurones zibonisa ukuba iipropati zangaphakathi ze-ion channels zingachaza, ubuncinci, i-threshold yomatshini kunye ne-kinetics yokulungelelanisa i-mechanoreceptors echazwe kumashumi eminyaka 1960. 80 usebenzisa amalungiselelo e-ex vivo.
IiProteni eziPhucukayo zeMechanosensitive
Imisinga ye-ion ye-Mechanosensitive kwi-neurons ye-somatosensory ibonakaliswe kakuhle, ngokuchaseneyo, incinci iyaziwa malunga nobunikazi beemolekyuli ezidibanisa i-mechanotransduction kwizilwanyana ezincelisayo. Izikrini ze-Genetic kwi-Drosophila kunye ne-C. elegans ziye zachonga iimolekyuli ze-mechanotransduction zabaviwa, kuquka i-TRP kunye ne-degenerin / epithelial Na + channel (Deg / ENaC) iintsapho.73 Imizamo yakutshanje yokucacisa isiseko se-molecular ye-mechanotransduction kwizilwanyana ezincelisayo ziye zagxila kakhulu kwi-homologs yalaba baviwa. . Ukongezelela, abaninzi balaba baviwa bakhona kwi-cutaneous mechanoreceptors kunye ne-somatosensory neurones (Umfanekiso 2).
I-Acid-Sensing Ion Channels
I-ASICs iyingxenye ye-proton-gated subgroup ye-degenerin�epithelial Na + yentsapho yesiteshi.74 Amalungu amathathu osapho lwe-ASIC (ASIC1, ASIC2 kunye ne-ASIC3) abonakaliswe kwi-mechanoreceptors kunye ne-nociceptors. Indima yeziteshi ze-ASIC iphandwe kwizifundo zokuziphatha kusetyenziswa iigundane ngokucima okujoliswe kuyo kwiijene ze-ASIC. Ukususwa kwe-ASIC1 ayitshintshi umsebenzi we-cutaneous mechanoreceptors kodwa kwandisa uvakalelo lwe-mechanical of afferents innervating gut.75 I-ASIC2 iigundane zokunkqonkqoza zibonisa ukuncipha kovakalelo lokulungelelanisa ngokukhawuleza ii-LTMRs zesikhumba. zombini i-visceral mechano-nociception kunye ne-cutaneous mechanosensation.76 I-ASIC2 ukuphazamiseka kunciphisa uvakalelo lwe-mechano ye-visceral afferents kwaye kunciphisa iimpendulo ze-HTMR ze-cutaneous kwi-stimuli eyingozi.77
Ijelo leSamkelo elidlulayo
I-TRP superfamily ihlulwe kwiintsapho ezithandathu kwizilwanyana ezincelisayo.78 Phantse zonke i-subfamilies ze-TRP zinamalungu axhunywe kwi-mechanosensation kwiinkqubo zeeseli ezahlukeneyo.79 Kwi-mammalian sensory neurones, nangona kunjalo, iziteshi ze-TRP ziyaziwa kakhulu ngokuva ulwazi lwe-thermal kunye nokudibanisa ukuvuvukala kwe-neurogenic, kunye neziteshi ezimbini ze-TRP kuphela, i-TRPV4 kunye ne-TRPA1, ziye zabandakanyeka ekuphenduleni kokuthinta. Ukuphazamisa ukubonakaliswa kwe-TRPV4 kwiigundane kuneempembelelo ezithobekileyo kuphela kwimida ye-acute mechanosensory, kodwa kunciphisa kakhulu uvakalelo kwi-stimuli eyingozi ye-mechanical stimuli.80,81 TRPV4 yinto ebalulekileyo ekumiseni impendulo ye-nociceptive neurones kuxinzelelo lwe-osmotic kunye ne-hyperalgesia yomatshini ngexesha lokuvuvukala.82,83, I-1 TRPA1 ibonakala inendima kwi-hyperalgesia yomatshini. I-TRPA1-iimpuku ezinqongopheleyo zibonisa iintlungu ze-hypersensitivity. I-TRPA84,85 inegalelo ekutshintsheni i-mechanical, ibanda kunye ne-chemical stimuli kwi-nociceptor sensory neurones kodwa kubonakala ngathi akubalulekanga kwi-hair-cell transduction.XNUMX
Akukho bungqina bucacileyo obubonisa ukuba amajelo e-TRP kunye neeshaneli ze-ASIC ezichazwe kwizilwanyana ezanyisayo zifakwe kwisango ngomatshini. Akukho nanye kwezi tshaneli ezibonakaliswe nge-heterologously recapitulate utyikityo lombane lwemisinga ye-mechanosensitive ebonwa kwindawo yabo yendalo. Oku akuthinteli ukuba kunokwenzeka ukuba i-ASICs kunye ne-TRPs iziteshi ziyi-mechanotransducers, zinikwe ukungaqiniseki ukuba ingaba ishaneli ye-mechanotransduction inokusebenza ngaphandle komxholo weselula (jonga icandelo kwi-SLP3).
Iiprotheyini zePiezo
I-Piezo protiens isanda kuchongwa njengabaviwa abathembisayo kwiiprotheni ze-mechanosensing yi-Coste kunye nabasebenzisana nabo. . I-Piezo 86,87 ininzi kwii-DRGs, ngelixa i-Piezo 1 ibonakala ngokungaqhelekanga. I-Piezo-induced mechanosensitive currents inhibited yi-gadolinium, i-ruthenium ebomvu kunye ne-GsMTx2 (i-toxin evela kwi-tarantula Grammostola spatulata) kune-Piezo 38. Ngokufana nemisinga ye-endogenous mechanosensitive currents, ii-currents ezixhomekeke kwi-Piezo zinamandla okubuyisela umva malunga ne-38 mV kwaye i-cation ayikhethi, kunye ne-Na+, K+, Ca2+ kunye ne-Mg1+ zonke zingena kumjelo ongaphantsi. Ngokufanayo, i-piezo-dependent currents ilawulwa yi-membrane enokubakho, kunye nokucotha okuphawulekayo kwe-kinetics yangoku kwi-depolarized potentials.4
Iiprotheyini ze-Piezo ngokungathandabuzekiyo ziyi-protein ze-mechanosensing kwaye zabelana ngeempawu ezininzi zokulungelelanisa ngokukhawuleza imisinga ye-mechanosensitive kwi-sensory neurones. Unyango lwee-neurones ze-DRG ezikhulisiwe kunye ne-Piezo 2 i-RNA emfutshane ephazamisayo iyancipha umlinganiselo we-neurones ngokukhawuleza ukulungelelanisa ngoku kunye nokunciphisa ipesenti ye-mechanosensitive neurones.86 Imimandla ye-Transmembrane ifumaneka kuzo zonke iiprotheni ze-piezo kodwa akukho mboniso ocacileyo we-pore-containing motifs okanye i-ion channel signatures ziye zafunyanwa. ichongiwe. Nangona kunjalo, i-mouse i-Piezo iprotheni ye-1 ihlanjululwe kwaye ihlaziywe kwi-asymmetric lipid bilayers kunye ne-liposome iifom ze-ion channels ezithintekayo kwi-ruthenium ebomvu.87 Isinyathelo esibalulekileyo ekuqinisekiseni i-mechanotransduction ngokusebenzisa iziteshi ze-Piezo kukusebenzisa iindlela ze-vivo ukugqiba ukubaluleka kokusebenza ekuboniseni umqondiso. Ulwazi lunikezwe kwi-Drosophila apho ukucima ilungu elilodwa le-Piezo linciphisa impendulo yomatshini kwi-stimuli enobungozi, ngaphandle kokuchaphazela ukubamba okuqhelekileyo.89 Nangona isakhiwo sabo sihlala sizimisele, le ntsapho yencwadana yeeprotheni ze-mechanosensitive yinto ethembisayo yophando lwexesha elizayo, ngaphaya komda. yemvakalelo yokuchukumisa. Ngokomzekelo, uphando olutshanje kwizigulane ezine-anemia (i-xerocytosis ye-hereditary) ibonisa indima ye-Piezo 1 ekugcineni i-erythrocyte volume homeostasis.90
Ijelo elifana neTransmembrane (TMC)
Uphononongo olutshanje lubonisa ukuba iiprotheni ezimbini, i-TMC1 kunye ne-TMC2, ziyimfuneko kwi-hair cell mechanotransduction.91 I-Hereditary deafness ngenxa ye-TMC1 yokuguqulwa kofuzo kwabikwa kubantu kunye neegundane.92,93 Ubukho bala majelo ayikabonakali kwinkqubo ye-somatosensory. , kodwa kubonakala ngathi isikhokelo esihle sokuphanda.
I-Stomatin-NjengeProtein 3 (SLP3)
Ukongezelela kwiitshaneli zokutshintshwa, ezinye iiprotheyini ze-accessory ezidityaniswe nomjelo zibonakaliswe ukuba zidlala indima ekuchukumiseni ukuchukumisa. I-SLP3 ibonakaliswe kwii-neuron ze-DRG ezincancisayo. Izifundo ezisebenzisa iigundane eziguqukayo ezingenayo i-SLP3 zibonise utshintsho kwi-mechanosensation kunye nemisinga ye-mechanosentive.94,95 Umsebenzi ochanekileyo we-SLP3 uhlala ungaziwa. Isenokuba ngumnxibelelanisi phakathi kweshaneli ye-mechanosensitive kunye ne-microtubules engaphantsi, njengoko kucetywayo kwi-C. elegans homolog MEC2.96 Kutshanje GR. I-lab ye-Lewin iphakamise ukuba i-tether yenziwe yi-DRG sensory neurones kwaye idibanisa i-ion channel ye-mechanosensitive kwi-matrix engaphandle. I-RA-mechanosensitive currents iphinda ivinjwe yi-lamin-97, iprotheni ye-matrix eveliswa yi-keratinocytes, iqinisa i-hypothesis ye-modulation ye-mechanosensitive yangoku ngama-protein angaphandle.332
I-K+ Channel Intsapho engaphantsi
Ngokuhambelana ne-cationic depolarizing misinga ye-mechanosensitive, ubukho be-repolarizing mechanosensitive K+ currents buphantsi kophando. Iziteshi ze-K + kwiiseli ze-mechanosensitive zinokunyathela kwibhalansi yangoku kwaye zibe negalelo ekuchazeni umda womatshini kunye nexesha lokulungelelanisa i-mechanoreceptors.
Amalungu e-KCNK angamalungu e-domain ye-pore ye-K + channel (K2P) yentsapho.99,100 I-K2P ibonisa uluhlu oluphawulekayo lokulawulwa ngamaselula, ama-agent angokwenyama kunye ne-pharmacological, kubandakanywa utshintsho lwe-pH, ukushisa, ukunweba kunye ne-membrane deformation. Ezi K2P ziyasebenza ekuphumleni inwebu enokubakho. Iinqununu ezininzi ze-KCNK zibonakaliswe kwii-neurones ze-somatosensory.101 KCNK2 (TREK-1), i-KCNK4 (TRAAK) kunye ne-TREK-2 iziteshi ziphakathi kweendlela ezimbalwa apho i-mechanical gating ngqo nge-membrane stretch iboniswe.102,103
Iimpuku ezinofuzo oluphazamisekileyo lwe-KCNK2 lubonise ubuntununtunu obuphuculweyo bobushushu kunye ne-stimuli ethambileyo yomatshini kodwa umda oqhelekileyo wokurhoxisa kuxinzelelo oluyingozi lomatshini olusetyenziswa kwi-hindpaw usebenzisa uvavanyo lwe-Randall�Selitto. iimeko. I-KCNK104 iigundane zokunkqonkqoza zazine-hypersensitive kwi-mild stimulation mechanical, kwaye le hypersensitivity yandiswe ngokungasebenzi okongeziweyo kwe-KCNK2 Ukonyuka kwe-mechanosensitivity kwezi mpuku ze-knockout kunokuthetha ukuba ukolula ngokuqhelekileyo kusebenze zombini i-depolarizing kunye ne-repolarizing currents ze-mechanosensitive ngendlela elungelelanisiweyo, ngokufanayo nokungalingani. ukudiliza kunye nokuphinda kuguqulwe imisinga yamandla ombane.
I-KCNK18 (i-TRSK) inegalelo elikhulu kwimvelaphi ye-K + yokuziphatha elawula ukuphumla kwe-membrane ye-somatosensory neurones.106 Nangona ingaziwa ukuba i-KCNK18 ivakalelwa ngokuthe ngqo kwi-mechanical stimulation, inokudlala indima ekuphenduleni iimpendulo kwi-touch light, kunye ne-stimuli ebuhlungu yomatshini. I-KCNK18 kunye ne-KCNK3 encinci, kucetywayo ukuba ibe yithagethi ye-molecular ye-hydroxy-?-sanshool, i-compound efunyenwe kwi-peppercorns ye-Schezuan eyenza i-receptors ye-touch kwaye ibangele ukuvakalelwa kwabantu.107,108.
I-voltage exhomekeke kwi-K + channel ye-KCNQ4 (Kv7.4) ibalulekile ekusetheni isantya kunye nokukhethwa kwefrikhwensi ye-subpopulation yokulungelelanisa ngokukhawuleza i-mechanoreceptors kuzo zombini iimpuku kunye nabantu. Uguqulo lwe-KCNQ4 luye lwanxulunyaniswa nohlobo lokungeva oluzuzwa lufuzo. Okubangela umdla uphononongo lwakutsha nje lubeka i-KCNQ4 kwi-peripheral nerve endings ye-cutaneous elungisa ngokukhawuleza i-follicle yeenwele kunye ne-Meissner corpuscle. Ngokufanelekileyo, ukulahleka komsebenzi we-KCNQ4 kukhokelela ekuphuculeni okukhethiweyo kwe-mechanoreceptor sensitivity kwi-vibration ephantsi-frequency. Ngokucacileyo, abantu abanokulahlekelwa kokuva emva kwexesha ngenxa yokuguqulwa kwezinto eziphambili ze-KCNQ4 gene zibonisa ukusebenza okuphuculweyo ekuboneni i-amplitude encinci, i-low-frequency vibration.109

Insight of Dr. Alex Jimenez
Ukuchukumisa kuthathwa njengenye yezona zivamvo zintsonkothileyo emzimbeni womntu, ngakumbi ngenxa yokuba akukho lungu lithile eliphetheyo. Kunoko, ingqiqo yokuchukumisa iyenzeka ngokusebenzisa i-receptors ye-sensory, eyaziwa ngokuba yi-mechanoreceptors, efumaneka kulo lonke ulusu kwaye iphendule kuxinzelelo lomatshini okanye ukuphazamiseka. Kukho iintlobo ezine eziphambili ze-mechanoreceptors kwi-glabrous, okanye inwele, isikhumba sezilwanyana ezincelisayo: i-lamellar corpuscles, i-tactile corpuscles, i-Merkel nerve endings kunye ne-bulbous corpuscles. I-Mechanoreceptors isebenze ukwenzela ukuvumela ukufunyanwa kwe-touch, ukwenzela ukubeka iliso kwindawo yemisipha, amathambo kunye namalungu, aziwa ngokuba yi-proprioception, kunye nokubona izandi kunye nokuhamba komzimba. Ukuqonda iindlela zesakhiwo kunye nomsebenzi wala ma-mechanoreceptors yinto ebalulekileyo ekusebenziseni unyango kunye nonyango lokulawula intlungu.
isiphelo
Ukuchukumisa yimvakalelo entsonkothileyo kuba imele iimpawu ezahlukeneyo zokuchukumisa, ezizezi, ukungcangcazela, ukumila, ukuthungwa, ulonwabo kunye neentlungu, kunye nemidlalo eyahlukeneyo ecalulayo. Ukuza kuthi ga ngoku, imbalelwano phakathi kwe-touch-organ kunye ne-psychophysical sense yayinxibelelene kwaye iimpawu zemolekyuli ezikhethekileyo ziyavela. Uphuhliso lovavanyo lwempuku oluhambelana nokwahluka kokuziphatha ngokuchukumisa kuyafuneka ngoku ukulungiselela ukuchongwa kwegenomics kwixesha elizayo. Ukusetyenziswa kweegundane ezingenayo i-subsets ethile yeentlobo ze-sensory afferent ziya kuququzelela kakhulu ukuchongwa kwe-mechanoreceptors kunye ne-sensory afferent fibers ezinxulumene ne-touch modality ethile. Kuyathakazelisa ukuba iphepha lamva nje livula umbuzo obalulekileyo wesiseko sofuzo seempawu ze-mechanosensory kumntu kwaye ubonisa ukuba ukuguqulwa kwemfuza enye kunokuchaphazela kakubi uvakalelo lokuthintela. inkqubela ngokuchonga ngokuchanekileyo i-subset ye-sensory neurones edityaniswe kwindlela yokuchukumisa okanye ukusilela kokuchukumisa.
Ukubuyisela, inkqubela yenziwe ukucacisa iimpawu ze-biophysical ze-mechano-gated currents.64 Ukuphuhliswa kweendlela ezintsha kwiminyaka yamuva nje, ukuvumela ukubeka iliso kwi-membrane ye-tension tension, ngelixa urekhoda i-mechano-gated yangoku, ibonakalise indlela ebalulekileyo yokulinga ukuchaza. i-mechanosensitive currents kunye nokulungelelaniswa okukhawulezileyo, okuphakathi kunye nokucothayo (kuhlaziywa kwi-Delmas kunye nabasebenzisana nabo) .66,111 Ixesha elizayo liya kuba libonisa indima yezakhiwo ezikhoyo kwiindlela zokulungelelanisa i-mechanoreceptors esebenzayo kunye negalelo le-mechanosensitive K + currents kwi-excitability of Ii-LTMR kunye nee-HTMRs.
Ubume bemolekyuli bemisinga ye-mechano-gated kwizilwanyana ezanyisayo kwakhona sisihloko sophando esithembisayo. Uphando lwexesha elizayo luya kuqhubela phambili kwiimbono ezimbini, okokuqala ukugqiba indima ye-molecule ye-accessory ukuba i-tether channels kwi-cytoskeleton kwaye iya kufuneka inike okanye ilawule i-mechanosensitivity ye-ion channels ezifana ne-TRP kunye ne-ASIC / iintsapho ze-EnaC. Okwesibini, ukuphanda indawo enkulu kunye nethembisayo yegalelo leetshaneli zePiezo ngokuphendula imibuzo ephambili, ngokumalunga ne-permeation kunye neendlela zokungena, i-subset ye-sensory neurones kunye neendlela zokuthintela ezibandakanya i-Piezo kunye nendima yePiezo kwiiseli ezingekho neuronal ezinxulumene nazo. I-mechanosensation.
Umqondo wokuchukumisa, xa kuthelekiswa nokubona, ukunambitha, isandi kunye nephunga, ezisebenzisa izitho ezithile ukucubungula ezi mvakalelo, zingenzeka kuwo wonke umzimba ngokusebenzisa i-receptors encinci eyaziwa ngokuba yi-mechanoreceptors. Iindidi ezahlukeneyo ze-mechanoreceptors zinokufumaneka kwiindawo ezahlukeneyo zesikhumba, apho ziyakwazi ukubona uluhlu olubanzi lokuvuselela oomatshini. Eli nqaku lingasentla lichaza izinto ezibalaseleyo ezibonisa inkqubela phambili yezakhiwo kunye neendlela zokusebenza ze-mechanoreceptors ezinxulumene nomqondo wokuchukumisa. Ulwazi olubhekiselele kwiZiko leSizwe loLwazi lweBiotechnology (NCBI). Ububanzi bolwazi lwethu bukhawulelwe kwi-chiropractic kunye nokulimala komgogodla kunye neemeko. Ukuxoxa ngalo mbandela, nceda uzive ukhululekile ukubuza uGqr. Jimenez okanye uqhagamshelane nathi apha915-850-0900 .
Ikhutshwe nguDkt. Alex Jimenez

Imixholo eyongezelelweyo: Ubunzima bokubuyisela
Umqolo obuhlungu enye yezona zizathu ezibangeleko zokukhubazeka kunye neentsuku eziphosakeleyo emhlabeni wonke. Njengokuba kunjalo, iintlungu zenziwa emva kweyona sibini isizathu esivakalayo sokutyelela iofisi yee-dkt. Ngokumalunga neepesenti ze-80 zabemi ziya kuba nolunye uhlobo lweentlungu zentlungu ubuncinane kanye kanye ebomini babo. Umgudu uyisakhiwo esiyinkimbinkimbi esakhiwa ngamathambo, amajoyina, iigaments kunye nezihlunu, phakathi kwezinye iifomthi ezithambileyo. Ngenxa yoko, ukulimala kunye / okanye iimeko ezihlaziyiweyo, ezifana disni, ekugqibeleni unokukhokelela kwiimpawu zentlungu. Ukulimala kwezemidlalo okanye ukulimala kwengozi yeemoto ngokuqhelekileyo kubangelwa yintlungu yokubuhlungu, kodwa ngamanye amaxesha ukuhamba kwezinto ezilula kunokuba neziphumo ezibuhlungu. Ngethamsanqa, ezinye iindlela zokhathalela unyango, ezifana nokunyamekelwa kwe-chiropractic, kunokunceda ukubuyisela intlungu emva kokusetyenziswa kwemilenze kunye nokunyanzeliswa kwemigaqo, ekugqibeleni ukuphucula intlungu.

ISIHLOKO ESIBALULEKILEYO: Ulawulo oluPhantsi lwePain Pain
IINKCUKACHA NGOKUGQITHISILEYO: UKONGEZA KWEZINTLOKO: PainUbuhlungu obungapheliyo kunye nonyango
Ngenanto
Ucaphulo
Vala i-Accordion
Umgangatho wobuchule wokuSebenza *
Ulwazi olulapha ku "Iindlela zokuSebenza kunye neziSebenzi zeMechanoreceptors"Akujoliswanga ukuthatha indawo yobudlelwane obubodwa kunye nomntu oqeqeshiweyo wezempilo okanye ugqirha onelayisensi kwaye akusiyo isiluleko sonyango. Sikhuthaza ukuba wenze izigqibo zezempilo ngokusekelwe kuphando lwakho kunye nentsebenziswano kunye nochwepheshe bezempilo abaqeqeshiweyo.
Ulwazi lweBlog kunye neengxoxo zoMda
Umda wethu wolwazi ilinganiselwe kwiChiropractic, i-musculoskeletal, amayeza omzimba, impilo, igalelo le-etiological ukuphazamiseka kwe-viscerosomatic ngaphakathi kweentetho zeklinikhi, ezinxulumene ne-somatovisceral reflex clinical dynamics, i-subluxation complexes, imiba yezempilo ebuthathaka, kunye / okanye amanqaku amayeza asebenzayo, izihloko kunye neengxoxo.
Sibonelela kwaye sibonise intsebenziswano yeklinikhi neengcaphephe kumacandelo ahlukeneyo. Ingcali nganye ilawulwa ngumsebenzi wabo wobugcisa kunye negunya labo lokufumana iphepha-mvume. Sisebenzisa iiprothokholi ezisebenzayo zempilo kunye nempilo entle ukunyanga nokuxhasa ukhathalelo lokwenzakala okanye ukuphazamiseka kwenkqubo ye-musculoskeletal.
Iividiyo zethu, izithuba, izihloko, imixholo, kunye nokuqonda zibandakanya imiba yezonyango, imiba, kunye nezihloko eziyelelene kwaye zixhase ngokuthe ngqo okanye ngokungathanga ngqo umda wokuziqhelanisa wethu.
I-ofisi yethu izamile ngokufanelekileyo ukubonelela ngeengcaphulo ezixhasayo kwaye ichonge uphando olufanelekileyo lophando okanye izifundo ezixhasa izithuba zethu. Sinikezela ngeekopi zophando ezixhasayo ezifumanekayo kwiibhodi ezilawulayo nakuluntu ngesicelo.
Siyaqonda ukuba sigubungela imicimbi efuna inkcazo eyongezelelweyo yokuba inganceda njani kwisicwangciso esithile sokhathalelo okanye inkqubo yonyango; ke, ukuqhubeka nokuxoxa ngombandela ongentla, nceda ukhululeke ukubuza UDkt Alex Jimenez, DC, okanye qha ga mshelana nathi 915-850-0900.
Silapha ukunceda wena kunye nosapho lwakho.
Iintsikelelo
UDkt Alex Jimenez D.C., I-MSACP, RN*, I-CCST, IFMCP*, I-CIFM*, I-ATN*
email: qeqeshi@elpasofunctionalmedicine.com
Ilayisenisi njengoGqirha weChiropractic (DC) kwi Texas & New Mexico*
Texas DC Ilayisensi # TX5807, New Mexico DC Ilayisensi # I-NM-DC2182
Unikwe Ilayisensi njengoMongikazi oBhalisiweyo (RN*) in Florida
Florida License RN Ilayisensi # I-RN9617241 (Nombolo yolawulo. 3558029)
Ubume obubambeneyo: ILayisensi yeeNkcazo ezininzi: Ugunyaziswe Ukuziqhelanisa I-40 States*
UGqr. Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Ikhadi lam loShishino lweDijithali





